 آرمان مهدویت مذهبی ، اعتقادی
آرمان مهدویت مذهبی ، اعتقادی
این سوالات، ابهامات و پیچیدگی ها، موجب شده است تا بعد از فراگیر شدن استفاده از کامپیوتر و نرم افزارهای کامپیوتری، زیست شناسان و تکامل شناسان از نرم افزارها و برنامه های کامپیوتری متعددی برای تلفیق و مرتبط کردن الگوریتم های ناهمگون بهره ببیند تا شاید بتوانند مشکلات ناشی از ناهماهنگی ها و تناقضات مشاهده شده در آنالیز ژن های مختلف را به نحوی کاهش دهند!
بسم الله الرحمن الرحیم
ادامه ی مبحث فسیل های « نئاندرتال ها » و مطالعات ژنتیک
G) علاوه بر مطالبی که تاکنون پیرامون مشکلات مربوط به انواع مطالعات ژنتیکی انجام شده روی فسیل ها ذکر گردید، مشکلات جدی نیز در مورد مطالعات مبتنی بر پروتکل « DNA باستانی : Ancient DNA » استفاده کننده از DNA میتوکندریال (mtDNA) وجود دارد.
DNA میتوکندریال (mtDNA)، نقش مهمی در مطالعات مبتنی بر روش « DNA باستانی : Ancient DNA » بازی می کند و بسیاری از آنالیزهای انجام شده بر روی DNA فسیل ها، با استفاده از DNA میتوکندریال (mtDNA) صورت گرفته است و فسیل های « نئاندرتال ها » نیز از آن بی نصیب نبوده اند!(۱۹۴)
DNA میتوکندریال (mtDNA)، نقش مهمی در مطالعات مبتنی بر روش « DNA باستانی : Ancient DNA » دارد و بسیاری از آنالیزهای انجام شده بر روی DNA فسیل ها، با استفاده از DNA میتوکندریال (mtDNA) صورت گرفته است و فسیل های « نئاندرتال ها » نیز از آن بی نصیب نبوده اند!
DNA میتوکندریال (mtDNA)، نقش مهمی در مطالعات مبتنی بر روش « DNA باستانی : Ancient DNA » دارد و بسیاری از آنالیزهای انجام شده بر روی DNA فسیل ها، با استفاده از DNA میتوکندریال (mtDNA) صورت گرفته است و فسیل های « نئاندرتال ها » نیز از آن بی نصیب نبوده اند!
استفاده از DNA میتوکندریال (mtDNA) در مطالعات فسیلی مبتنی بر روش « DNA باستانی : Ancient DNA »، به چند دلیل مورد توجه بوده است: اولاً گرچه DNA میتوکندریال (mtDNA)، درصد بسیار کمی از کل DNA سلولی را تشکیل می دهد، اما همین درصد کم، در کپی های متعدد و فراوان در سطح سلول ها پخش شده اند(۱۷۰) و بازیابی آن ها امکان پذیر است. ثانیاً به دلیل این که مطابق مفروضات علمی (تا سال ۱۹۹۵ میلادی)، انتقال DNA میتوکندریال (mtDNA)، از مادر به فرزندان در طی نسل ها رخ داده و « توارث مادری » دارد(۱۷۳)، گزینه ی جذابی برای مطالعات تبارشناسی و از جمله بررسی زمان جدا شدن نسل ها از یکدیگر به شمار می رود.(۱۷۳)
بنا بر مفروضات قبلی، DNA میتوکندریال (mtDNA) عمدتاً تنها از مادران به فرزندان انتقال می یابد و به همین ترتیب، عمدتاً تنها دختران این مادران می توانند DNA میتوکندریال (mtDNA) کسب شده را به فرزندان خود انتقال دهند و … بدین ترتیب، عمدتاً تنها مادران و دختران می توانند عامل انتقال نسل به نسل DNA میتوکندریال به نسل بعد از خود باشند! به همین دلیل توارث DNA میتوکندریال (mtDNA) را « توارث مادری » می نامند که می تواند تنها از طریق مادران به دختران و سپس نواده های دختری و … انتقال یابد. با توجه به این مفروضات، DNA میتوکندریال (mtDNA) به عنوان یک گزینه ی جذاب برای و از جمله بررسی زمان جدا شدن نسل ها از یکدیگر به شمار می رود.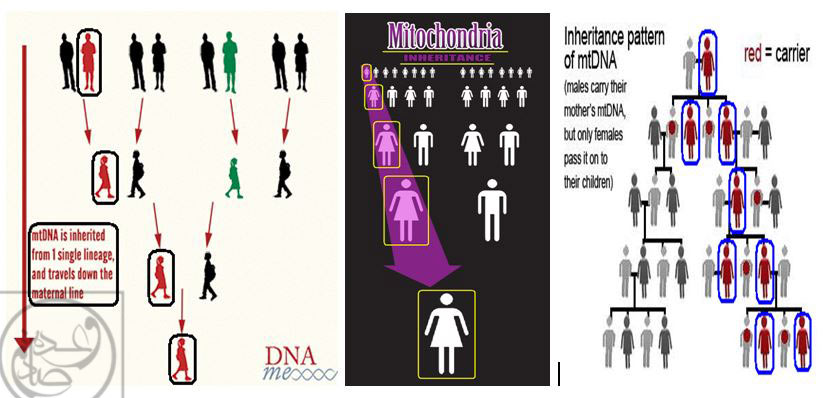
البته امروزه، کم کم مطالعه بر روی DNA هسته ای فسیل ها نیز رونق گرفته و این نوع مطالعات در حال افزایش است!(۱۹۵)
تا این جای کار برای مخاطبان محترم و به خصوص عزیزانی که مطالعاتی در حوزه ی « زیست شناسی » دارند، مسئله ی عجیب و غریبی نیست! اما موضوع هنگامی جالب می شود که بدانیم، مفروضات قبلی در مورد DNA میتوکندریال (mtDNA) از حوالی سال ۱۹۹۵ میلادی به بعد، با ابهامات و ایرادات جدی موجه شده است که برخی از آن ها به شرح زیر می باشند:
الف) برخلاف تصورات قبلی در مورد این که DNA میتوکندریال (mtDNA) صرفاً به صورت توارث مادری انتقال می یابد، شواهدی به دست آمده است که نشان می دهد در برخی موارد، DNA میتوکندریال (mtDNA) موجود در اسپرم پدر نیز بعد از لقاح می تواند زنده مانده و به فرزندان انتقال یابد! (۱۹۶)
برخلاف تصورات قبلی در مورد این که DNA میتوکندریال (mtDNA) صرفاً به صورت توارث مادری انتقال می یابد، شواهدی به دست آمده است که نشان می دهد در برخی موارد، DNA میتوکندریال (mtDNA) موجود در اسپرم پدر نیز بعد از لقاح می تواند زنده مانده و به فرزندان انتقال یابد!
برخلاف تصورات قبلی در مورد این که DNA میتوکندریال (mtDNA) صرفاً به صورت توارث مادری انتقال می یابد، شواهدی به دست آمده است که نشان می دهد در برخی موارد، DNA میتوکندریال (mtDNA) موجود در اسپرم پدر نیز بعد از لقاح می تواند زنده مانده و به فرزندان انتقال یابد!
بدین ترتیب، مفروضات قبلی در مورد این که توارث DNA میتوکندریال (mtDNA) صرفاً به صورت « توارث مادری » است، زیر سوال می رود و احتمال « توارث پدری » یا « توارث توام » نیز در این میان مطرح می گردد!
ب) نکته ی دیگری که مفروضات قبلی را درباره ی DNA میتوکندریال (mtDNA) زیر سوال می برد، کشف مسئله ی تنوع داخل سلولی میتوکندری ها یا « هتروپلاسمی (ناهمگونی) : Heteroplasmy » است.
به عبارت دیگر، قبلاً تصور می شد که تمامی میتوکندری های داخل سلولی، کپی یکسانی از DNA دارند؛ حال آن که در مطالعات بعدی مشخص گردید که در بسیاری از موجودات زنده و از جمله انسان ها، ممکن است میتوکندری های مختلف داخل یک سلول، DNA های متفاوتی داشته باشند!:(۱۹۷)
تصویر راست، نمایی از « هوموپلاسمی : Homoplasmy » را نشان می دهد که در آن تمامی میتوکندری ها (بیضی های زرد رنگ)، حاوی DNA یکسانی هستند. قبلاً تصور می شد که تمامی سلول های بدن موجودات زنده از جمله انسان، دارای وضعیت « هوموپلاسمی : Homoplasmy » هستند. تصاویر چپ، نمایی از « هتروپلاسمی (ناهمگونی) : Heteroplasmy » زا نشان می دهند که در آن ها، میتوکندری های سلول (بیضی های زرد رنگ وقرمز رنگ)، حاوی DNA های متفاوتی هستند! امروزه مشخص گردیده است که پدیده ی « هتروپلاسمی (ناهمگونی) : Heteroplasmy »، یک پدیده ی واقعی و شایع در سلول های بدن موجودات زنده و از جمله انسان است!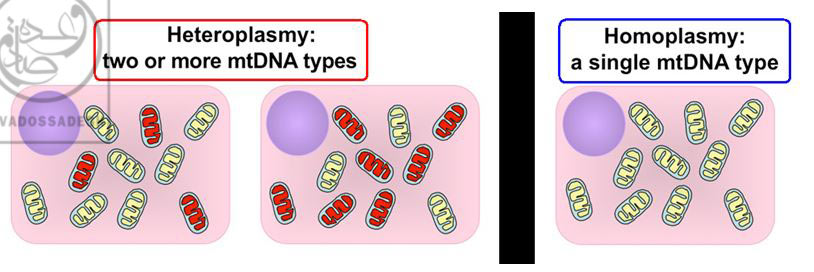
البته پدیده ی « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها، پدیده ی ناشایعی نیست! بلکه برخی مطالعات، شیوع « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها را در سلول های انسان ها، از ۱۰ درصد الی ۲۰ درصد افراد جوامع انسانی گزارش نموده اند:(۱۹۷)
پدیده ی « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها در داخل سلول ها، پدیده ی ناشایعی نیست! بلکه برخی مطالعات، شیوع « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها را در سلول های انسان ها، از ۱۰ درصد الی ۲۰ درصد گزارش نموده اند!
 اما نکته ی شگفت انگیز تر این که حتی در بدن یک انسان نیز، ممکن است از بافتی به بافت دیگر، میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها متغیر باشد!(۱۹۸) برای مثال، ممکن است میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها در سلول های عضلات اسکلتی، با میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها در سلول های پوست، متفاوت باشد! همچنین مشخص شده است که حتی با تغییر سن انسان نیز، ممکن است میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها تغییر کند!:(۱۹۸)
اما نکته ی شگفت انگیز تر این که حتی در بدن یک انسان نیز، ممکن است از بافتی به بافت دیگر، میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها متغیر باشد!(۱۹۸) برای مثال، ممکن است میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها در سلول های عضلات اسکلتی، با میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها در سلول های پوست، متفاوت باشد! همچنین مشخص شده است که حتی با تغییر سن انسان نیز، ممکن است میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها تغییر کند!:(۱۹۸)
 حتی در بدن یک انسان نیز، ممکن است از بافتی بهبافت دیگر، میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها متغیر باشد! برای مثال، ممکن است میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری هادر سلول های عضلات اسکلتی، با میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها در سلول های پوست، متفاوت باشد! همچنین مشخص شده است که حتی با تغییر سن انسان نیز، ممکن است میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها تغییر کند!
حتی در بدن یک انسان نیز، ممکن است از بافتی بهبافت دیگر، میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها متغیر باشد! برای مثال، ممکن است میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری هادر سلول های عضلات اسکلتی، با میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها در سلول های پوست، متفاوت باشد! همچنین مشخص شده است که حتی با تغییر سن انسان نیز، ممکن است میزان « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها تغییر کند!
بدین ترتیب با توجه به مطالعات انجام شده در طی سال های اخیر، مشخص گردیده است که مفروضات قبلی تکامل شناسان، در رابطه با « توارث DNA یکسان مادری » از مادر به فرزندان، نادرست بوده و موارد متعدد و اثبات شده ای از « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها در طی سال های اخیر کشف گردیده است! به نحوی که نه تنها در افراد یک خانواده، بلکه حتی در سلول های مختلف یک انسان و حتی در سنین مختلف عمر وی، میزان متغیر و گهگاه غیرقابل پیش بینی از « هتروپلاسمی (ناهمگونی) : Heteroplasmy » میتوکندری ها ملاحظه می گردد!!! با عنایت به این کشفیات، می توان گفت که مفروضات قبلی تکامل شناسان پیرامون « توارث مادری DNA میتوکندریال (mtDNA) »، با چالش ها و اشکالات جدی مواجه است!
ج) مسئله ی دیگری که در مقابل ادعاهای تکامل شناسان، چالش جدی ایجاد می نماید، این نکته است که برخلاف مفروضات قبلی تکامل شناسان که سرعت بروز جهش در DNA میتوکندریال (mtDNA) را طی نسل های مختلف، ثابت و یکسان فرض می کردند، امروزه مشخص شده است که سرعت جهش های ژنتیکی در « DNA میتوکندریال (mtDNA) »، متغیر و غیر قابل پیش بینی بوده و در برخی از موارد، سرعت بروز جهش ها در « DNA میتوکندریال (mtDNA) »، بسیار بیشتر از آن چیزی است که در محاسبات قبلی تکامل شناسان، لحاظ شده بود!(۱۹۹)
در واقع مطالعات جدید (پس از سال ۱۹۹۵ میلادی) نشان داده اند که جهش های ژنتیکی یا « موتاسیون : Mutation » ها(۱۷۲) در « DNA میتوکندریال (mtDNA) »، بسیار سریع تر از محاسبات قبلی، به وقوع می پیوندند!:(۱۹۹)
مسئله ی دیگری که در مقابل ادعاهای تکامل شناسان، چالش جدی ایجاد می نماید، این نکته است که برخلاف مفروضات قبلی تکامل شناسان که سرعت بروز جهش در DNA میتوکندریال (mtDNA) را طی نسل های مختلف، ثابت و یکسان فرض می کردند، امروزه مشخص شده است که سرعت جهش های ژنتیکی در « DNA میتوکندریال (mtDNA) »، متغیر و غیر قابل پیش بینی بوده و در برخی از موارد، سرعت بروز جهش ها در « DNA میتوکندریال (mtDNA) »، بسیار بیشتر از آن چیزی است که در محاسبات تکامل شناسان لحاظ شده بود! در واقع مطالعات جدید (پس از سال ۱۹۹۵ میلادی) نشان داده اند که جهش های ژنتیکی یا « موتاسیون : Mutation » ها در « DNA میتوکندریال (mtDNA) »، بسیار سریع تر از محاسبات قبلی، به وقوع می پیوندند!
مسئله ی دیگری که در مقابل ادعاهای تکامل شناسان، چالش جدی ایجاد می نماید، این نکته است که برخلاف مفروضات قبلی تکامل شناسان که سرعت بروز جهش در DNA میتوکندریال (mtDNA) را طی نسل های مختلف، ثابت و یکسان فرض می کردند، امروزه مشخص شده است که سرعت جهش های ژنتیکی در « DNA میتوکندریال (mtDNA) »، متغیر و غیر قابل پیش بینی بوده و در برخی از موارد، سرعت بروز جهش ها در « DNA میتوکندریال (mtDNA) »، بسیار بیشتر از آن چیزی است که در محاسبات تکامل شناسان لحاظ شده بود! در واقع مطالعات جدید (پس از سال ۱۹۹۵ میلادی) نشان داده اند که جهش های ژنتیکی یا « موتاسیون : Mutation » ها در « DNA میتوکندریال (mtDNA) »، بسیار سریع تر از محاسبات قبلی، به وقوع می پیوندند!
همان گونه که ملاحظه فرمودید، با توجه به کشفیات جدید علم ژنتیک، سرعت بروز جهش های ژنتیکی در « DNA میتوکندریال (mtDNA) » طی نسل های مختلف، متغیر بوده و عموماً جهش ها سریع تر از آن چیزی که قبلاً تصور می شد، به وقوع می پیوندند!(۱۹۹) این کشفیات، ادعاهای تکامل شناسان را که بر اساس مفروضات غلط قبلی شکل گرفته است، باطل می کند و محاسبات صورت گرفته توسط آنان را که بر اساس پیش فرض های قبلی انجام شده است، مبدل به کاغذپاره می نماید!
اما از مجموع مطالبی که در بخش های (الف)، (ب) و (ج) گفته شد، این نتیجه به دست می آید که ادعاهای تکامل شناسان در حوزه های تبارشناسی و تکامل، کاملاً سست و ضعیف بوده و بر اساس اطلاعات قبل از سال ۱۹۹۵ میلادی می باشد! (گر چه هنوز تکامل شناسان با بی شرمی کامل از آن ها در کتب و مقالاتشان استفاده می کنند و حتی اشاره ای به کشفیات جدید در این حوزه نمی نمایند!).
امروزه بسیاری از دانشمندان، این حقیقت را پذیرفته اند که مطالعه در حوزه ی « DNA میتوکندریال (mtDNA) »، بسیار پیچیده تر از آن است که قبلاً تصور می شد؛(۲۰۰) همچنین این محققان معتقدند که ادعاهای ساده انگارانه ی قبلی می بایست، تعدیل گردد و راهکارهای نوینی برای مطالعات جدید، در خدمت گرفته شود.(۲۰۰)
از سوی دیگر، تعدادی از محققین نیز به صراحت درباره ی عواقبی که این کشفیات جدید برای « تکامل شناسان » در پی دارد، اشاره کرده اند و این کشفیات را موجب نگرانی « تکامل شناسان » دانسته اند!:(۲۰۰)
امروزه بسیاری از دانشمندان، این حقیقت را پذیرفته اند که مطالعه در حوزه ی « DNA میتوکندریال (mtDNA) »، بسیار پیچیده تر از آن است که قبلاً تصور می شد؛ همچنین این محققان معتقدند که ادعاهای ساده انگارانه ی قبلی می بایست، تعدیل گردد و راهکارهای نوینی برای مطالعات جدید، در خدمت گرفته شود. از سوی دیگر، تعدادی از محققین نیز به صراحت درباره ی عواقبی که این کشفیات جدید برای « تکامل شناسان » در پی دارد، اشاره کرده اند و این کشفیات را موجب نگرانی « تکامل شناسان » دانسته اند!
امروزه بسیاری از دانشمندان، این حقیقت را پذیرفته اند که مطالعه در حوزه ی « DNA میتوکندریال (mtDNA) »، بسیار پیچیده تر از آن است که قبلاً تصور می شد؛ همچنین این محققان معتقدند که ادعاهای ساده انگارانه ی قبلی می بایست، تعدیل گردد و راهکارهای نوینی برای مطالعات جدید، در خدمت گرفته شود. از سوی دیگر، تعدادی از محققین نیز به صراحت درباره ی عواقبی که این کشفیات جدید برای « تکامل شناسان » در پی دارد، اشاره کرده اند و این کشفیات را موجب نگرانی « تکامل شناسان » دانسته اند!
امروزه بسیاری از دانشمندان، این حقیقت را پذیرفته اند که مطالعه در حوزه ی « DNA میتوکندریال (mtDNA) »، بسیار پیچیده تر از آن است که قبلاً تصور می شد؛ همچنین این محققان معتقدند که ادعاهای ساده انگارانه ی قبلی می بایست، تعدیل گردد و راهکارهای نوینی برای مطالعات جدید، در خدمت گرفته شود. از سوی دیگر، تعدادی از محققین نیز به صراحت درباره ی عواقبی که این کشفیات جدید برای « تکامل شناسان » در پی دارد، اشاره کرده اند و این کشفیات را موجب نگرانی « تکامل شناسان » دانسته اند!
امروزه بسیاری از دانشمندان، این حقیقت را پذیرفته اند که مطالعه در حوزه ی « DNA میتوکندریال (mtDNA) »، بسیار پیچیده تر از آن است که قبلاً تصور می شد؛ همچنین این محققان معتقدند که ادعاهای ساده انگارانه ی قبلی می بایست، تعدیل گردد و راهکارهای نوینی برای مطالعات جدید، در خدمت گرفته شود. از سوی دیگر، تعدادی از محققین نیز به صراحت درباره ی عواقبی که این کشفیات جدید برای « تکامل شناسان » در پی دارد، اشاره کرده اند و این کشفیات را موجب نگرانی « تکامل شناسان » دانسته اند!
بدین ترتیب همان گونه که ملاحظه فرمودید، ادعاهای تکامل شناسان پیرامون دقت بالای مطالعات مبتنی بر « DNA میتوکندریال (mtDNA) »، با ابهامات و چالش های جدی مواجه شده و با توجه به این که ۵۰ درصد یا حتی بیش از ۵۰ درصد مطالعات مبتنی بر روش « DNA باستانی : Ancient DNA » که تاکنون انجام شده اند، بر اساس « DNA میتوکندریال (mtDNA) » و با توجه به مفروضات قبلی، طرح ریزی شده اند، باید گفت که حداقل نیمی از مطالعات ژنتیکی مبتنی بر روش « DNA باستانی : Ancient DNA » که تاکنون انجام شده اند، با چالش ها، ابهامات و سوالات جدی مواجه هستند!
گرچه برخی مشکلات ذکر شده در مورد « DNA میتوکندریال (mtDNA) » همچون « هتروپلاسمی » در مورد DNA هسته ای وجود ندارد، اما سایر مشکلاتی که قبلاً در مورد مشکلات موجود در روش « DNA باستانی : Ancient DNA » ذکر شد – اعم از تغییرات شیمیایی DNA بعد از مرگ، آلودگی نمونه های DNA، استاندارد نبودن روش های مطالعه بر روی DNA باستانی، تعداد کم فسیل های مورد مطالعه، و ورود DNA ویروس به DNA بدن انسان و جانوران (که اتفاقاً اختصاص به DNA هسته ای دارد) -، هم در DNA میتوکندریال (mtDNA) و هم در DNA هسته ای به چشم می خورد و مطالعات مربوط به هر دو نوع DNA را به چالش می طلبد! بدین ترتیب هر دو نوع DNA در مطالعات مبتنی بر روش « DNA باستانی : Ancient DNA » با مشکلات جدی رو به رو هستند!
H) اما یکی دیگر از مشکلاتی که در مورد مطالعات مبتنی بر روش « DNA باستانی : Ancient DNA » وجود دارد، مسئله ی عدم تطابق یافته های حاصل از مطالعه بر روی ژن ها و قطعات مختلف DNA است! در واقع این مشکل که از یک سو، هم در مطالعات « DNA باستانی : Ancient DNA » و هم در مطالعات DNA موجودات امروزی وجود دارد و از سوی دیگر، هم مطالعات انجام شده بر روی DNA میتوکندریال (mtDNA) و هم مطالعات مربوط به DNA هسته ای را با چالش مواجه می کند، موجب بروز اشکالات، ابهامات و سردرگمی های بسیار می گردد!
قبل از بررسی این مشکل، می بایست توضیح مختصری پیرامون « درخت فیلوژنتیک : Phylogenetic tree »(۲۰۱)، « درخت اتصال – همسایگی : tree Neighbor joining »(۲۰۲) و « درخت بایزی : Bayesian tree »(۲۰۳) بدهیم؛ « درخت فیلوژنتیک : Phylogenetic tree »(۲۰۱)، « درخت اتصال – همسایگی : Neighbor joining »(۲۰۲) و « درخت بایزی : Bayesian tree »(۲۰۳)، یکی از شایع ترین متدها و روش هایی هستند که « تکامل شناسان » از آن ها برای نشان دادن رابطه ی خویشاوندی موجودات بهره می برند. بدین ترتیب که موجودات زنده ی مد نظر خود را در « درخت » هایی به نمایش می گذارند که هر شاخه ی این درخت ها، متعلق به یک « موجود زنده » می باشد. در این درخت ها، هر چه موجودات از نظر تکاملی به هم نزدیکتر باشند و هر چه خویشاوندی بیشتر و شباهت ظاهری (آناتومیک)، فیزیولوژیک یا ژنتیک بیشتری داشته باشند، در شاخه های نزدیک تری قرار می گیرند و هر چه از نظر تکاملی از یکدیگر دورتر باشند و هر چه خویشاوندی کمتر، شباهت ظاهری (آناتومیک)، فیزیولوژیک یا ژنتیک کمتری داشته و از همدیگر دورتر باشند، در شاخه های دورتری نسبت به یکدیگر قرار می گیرند (البته به زعم تکامل شناسان!):
تصاویری از « درخت فیلوژنتیک : Phylogenetic tree »، « درخت اتصال – همسایگی : Neighbor joining tree » و « درخت بایزی : Bayesian tree »؛ هر شاخه ی این درخت ها، متعلق به یک « موجود زنده » می باشد. در این درخت ها، هر چه موجودات از نظر تکاملی به هم نزدیکتر باشند و هر چه خویشاوندی بیشتر و شباهت ظاهری (آناتومیک)، فیزیولوژیک یا ژنتیک بیشتری داشته باشند، در شاخه های نزدیک تری قرار می گیرند (موجودات زنده ی داخل کادر قرمز رنگ) و هر چه از نظر تکاملی از یکدیگر دورتر باشند و هر چه خویشاوندی کمتر، شباهت ظاهری (آناتومیک)، فیزیولوژیک یا ژنتیک کمتری داشته و از همدیگر دورتر باشند – به زعم تکامل شناسان! -، در شاخه های دورتری نسبت به یکدیگر قرار می گیرند (فاصله ی بین موجودات زنده ی کادر قرمز رنگ و کادر آبی رنگ)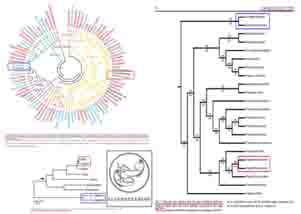
البته از نظر علمی، اختلافاتی بین تعاریف « درخت فیلوژنتیک : Phylogenetic tree »(۲۰۱)، « درخت اتصال – همسایگی : Neighbor joining tree »(۲۰۲) و « درخت بایزی : Bayesian tree »(۲۰۳) وجود دارد که از ذکر آن به دلیل جلوگیری از طولانی شدن بحث خودداری می نماییم؛ اما معمولاً « درخت فیلوژنتیک : Phylogenetic tree »(۲۰۱) برای مقاصد مقایسه ی ظاهری (آناتومیک) و ژنتیک کاربرد دارد، اما « درخت اتصال – همسایگی : Neighbor joining tree »(۲۰۲) و « درخت بایزی : Bayesian tree »(۲۰۳) بیشتر برای مقاصد مقایسه ی ژنتیکی یا ساختار پروتئینی و امثالهم کاربرد دارند.
به هر حال، آن چه اهمیت دارد، این است که نزدیکی یا دوری ظاهری (آناتومیک)، ژنتیکی، پروتئومیکی و … موجودات زنده (البته به زعم تکامل شناسان!) به وسیله ی درخت هایی همچون « درخت فیلوژنتیک : Phylogenetic tree »(۲۰۱)، « درخت اتصال – همسایگی : Neighbor joining tree »(۲۰۲) و « درخت بایزی : Bayesian tree »(۲۰۳) نمایش داده می شود که در این درختان، دوری یا نزدیکی شاخه ها ، معرف دوری یا نزدیکی موجودات زنده ی مورد نظر است.
قبلاً به مشکل عدم شباهت تقسیم بندی و درخت فیلوژنتیکی اسب های موسوم به اسب های ماقبل تاریخ بر اساس ظاهر فسیل های آن ها، با تقسیم بندی و درخت فیلوژنتیکی آن ها بر اساس آنالیز به روش « DNA باستانی : Ancient DNA » در بخش های قبلی همین مقاله اشاره کردیم.
اما مشکل هنگامی بیشتر خود را نشان می دهد که ملاحظه می گردد که مطالعات مختلف انجام شده بر روی بخش های مختلفی از DNA و بخش های مختلفی از ژن های گوناگون فسیل ها یا موجودات زنده ی کنونی، نتایج متفاوت و متغیری ارایه می دهند! برای مثال هنگامی که ۳ موجود زنده ی A، B و C از نظر ژنتیکی بررسی می گردند، نتایج آنالیز بر روی ژن (الف) این موجودات زنده، با نتایج آنالیز بر روی ژن (ب) این موجودات زنده، متفاوت می باشد!
تصاویر زیر، « درخت های فیلوژنتیک : Phylogenetic trees » چند گونه از « پلاسمودیوم : Plasmodium » ها را که انواع انسانی آن در انسان، بیماری « مالاریا » را ایجاد می کند، بر اساس ۳ پارامتر شامل « DNA هسته ای کد کننده ی زیر واحد کوچک RNA ریبوزومی (rRNA) موسوم به (SSU) »، « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » و نیز « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » نشان می دهد:(۲۰۴)
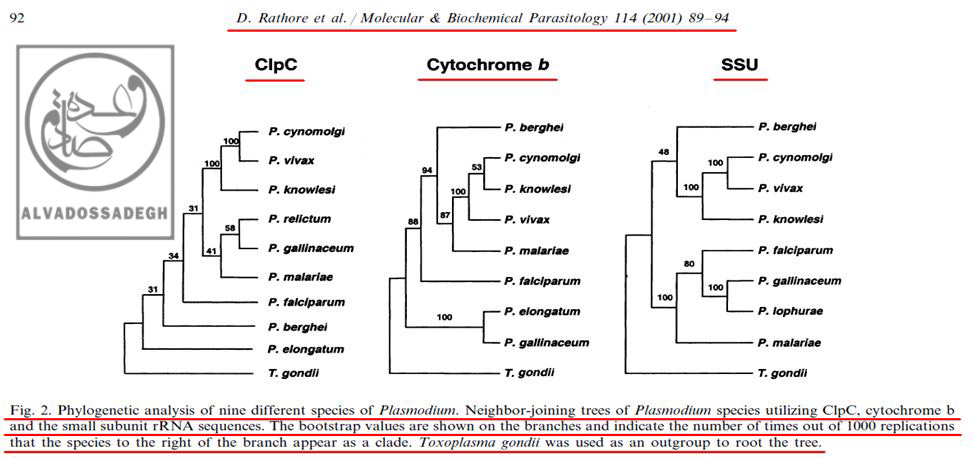 تصاویر فوق، « درخت های فیلوژنتیک : Phylogenetic trees » چند گونه از « پلاسمودیوم : Plasmodium » ها را که انواع انسانی آن در انسان بیماری « مالاریا » را ایجاد می کند، بر اساس ۳ پارامتر شامل « DNA هسته ای کد کننده ی زیر واحد کوچک RNA ریبوزومی (rRNA) موسوم به (SSU) »، « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » و نیز « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » نشان می دهد.
تصاویر فوق، « درخت های فیلوژنتیک : Phylogenetic trees » چند گونه از « پلاسمودیوم : Plasmodium » ها را که انواع انسانی آن در انسان بیماری « مالاریا » را ایجاد می کند، بر اساس ۳ پارامتر شامل « DNA هسته ای کد کننده ی زیر واحد کوچک RNA ریبوزومی (rRNA) موسوم به (SSU) »، « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » و نیز « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » نشان می دهد.
همان گونه که در تصاویر فوق مشاهده می فرمایید، علی رغم این که طبق ادعاهای تکامل شناسان، انتظار می رود که در مقایسه بین « گونه » های مختلف انگل « پلاسمودیوم »، آنالیز بخش های مختلف ژنی و DNAهای قسمت های مختلف سلول های آن ها (اعم از DNA هسته ای، DNA میتوکندریال (mtDNA) و DNA پلاستیدی)، درخت های فیلوژنتیک کم و بیش مشابهی به دست آید – تا همگام با هم سیر خویشاوندی و تکاملی مشابهی به نمایش بگذارند -، اما ملاحظه می گردد که آنالیز هر DNA، درخت فیلوژنتیک متفاوتی را به نمایش می گذارد!!!
برای مثال، در آنالیز به دست آمده از « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) »، مشاهده می شود که دو گونه ی « پلاسمودیوم الونگاتوم : P. elongatum » و « پلاسمودیوم گالیناسئوم : P. gallinaceum » قرابت زیادی با همدیگر از نظر این ژن دارند و بر روی دو شاخه ی مجاور قرار دارند، اما همین دو گونه یعنی « پلاسمودیوم الونگاتوم : P. elongatum » و « پلاسمودیوم گالیناسئوم : P. gallinaceum » هنگامی که از نظر « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » بررسی می گردند، تفاوت های فاحشی نشان می دهند و بر روی دو شاخه ی دور از هم قرار می گیرند!(۲۰۴)
مثال دیگر در همین آنالیز ها، مقایسه ی بین دو گونه ی « پلاسمودیوم گالیناسئوم : P. gallinaceum » و « پلاسمودیوم مالاریه : P. malariae » می باشد؛ به نحوی که آنالیز « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) »، جایگاه « پلاسمودیوم گالیناسئوم : P. gallinaceum » و « پلاسمودیوم مالاریه : P. malariae » را در شاخه های به نسبت نزدیک به هم و دارای قرابت زیاد نشان می دهد، اما آنالیز « DNA هسته ای کد کننده ی زیر واحد کوچک RNA ریبوزومی (rRNA) موسوم به (SSU) » و « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » برای همین دو گونه، جایگاه به مراتب دورتری را به نمایش می گذارد:(۲۰۴)
همان گونه که در تصاویر فوق مشاهده می فرمایید، علی رغم این که با توجه به ادعاهای تکامل شناسان، انتظار می رود که در مقایسه بین « گونه » های مختلف انگل « پلاسمودیوم »، آنالیز بخش های مختلف ژنی و DNAهای قسمت های مختلف سلول های آن ها (اعم از DNA هسته ای، DNA میتوکندریال (mtDNA) و DNA پلاستیدی)، درخت های فیلوژنتیک کم و بیش مشابهی به دست آید – تا همگام با هم سیر خویشاوندی و تکاملی مشابهی به نمایش بگذارند -، اما ملاحظه می گردد که آنالیز هر DNA، درخت فیلوژنتیک متفاوتی را به نمایش می گذارد!!! در آنالیز به دست آمده از « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) »، مشاهده می شود که دو گونه ی « پلاسمودیوم الونگاتوم : P. elongatum » (کادر قرمز رنگ) و « پلاسمودیوم گالیناسئوم : P. gallinaceum » (کادر سبز رنگ) قرابت زیادی با همدیگر از نظر این ژن دارند و بر روی دو شاخه ی مجاور قرار دارند، اما همین دو گونه یعنی « پلاسمودیوم الونگاتوم : P. elongatum » و « پلاسمودیوم گالیناسئوم : P. gallinaceum » هنگامی که از نظر « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » بررسی می گردند، تفاوت های فاحشی نشان می دهند و بر روی دو شاخه ی دور از هم قرار می گیرند! مثال دیگر در همین آنالیز ها مقایسه ی بین دو گونه ی « پلاسمودیوم گالیناسئوم : P. gallinaceum » (کادر سبز رنگ) و « پلاسمودیوم مالاریه : P. malariae » (کادر آبی رنگ) می باشد؛ به نحوی که آنالیز « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) »، جایگاه « پلاسمودیوم گالیناسئوم : P. gallinaceum » و « پلاسمودیوم مالاریه : P. malariae » را در شاخه های به نسبت نزدیک به هم و دارای قرابت زیاد نشان می دهد، اما آنالیز « DNA هسته ای کد کننده ی زیر واحد کوچک RNA ریبوزومی (rRNA) موسوم به (SSU) » و « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » برای همین دو گونه، جایگاه به مراتب دورتری را به نمایش می گذارد!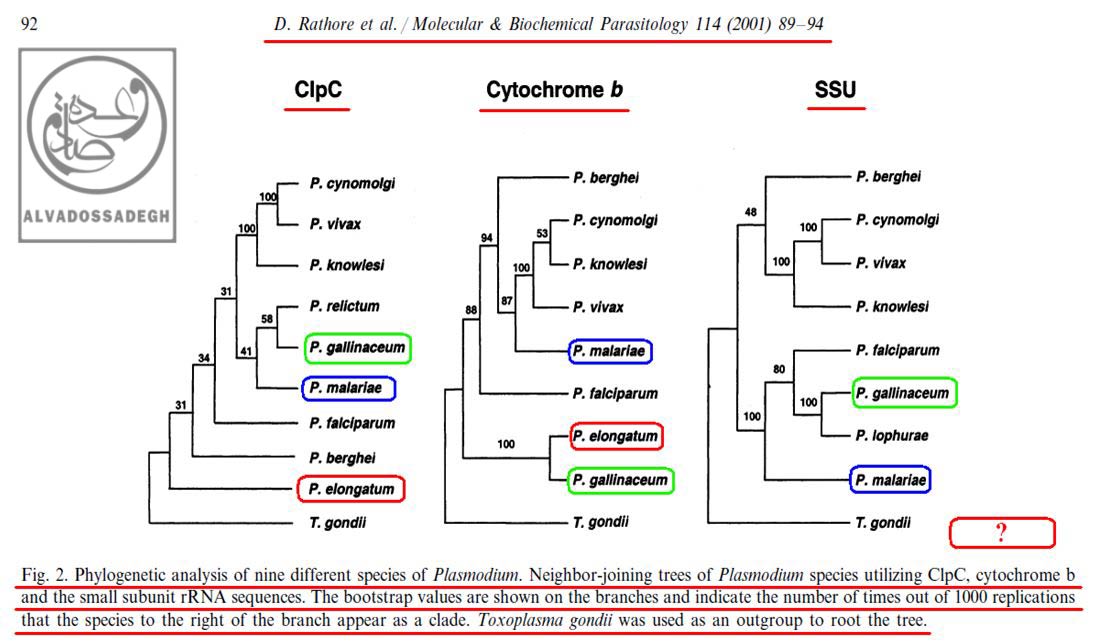
البته با دقت بیشتر در درخت های فیلوژنتیک ترسیم شده برای گونه های مختلف « پلاسمودیوم » ها، باز هم موارد زیادی از این گونه تناقضات و ناهماهنگی ها، مشخص می گردد! این مسئله نشان می دهد که بسته به این که کدام DNA سلولی مورد مطالعه واقع شود و حتی بسته به این که کدام قطعه از DNA یا کدام ژن بررسی گردد، درخت های فیلوژنتیک رسم شده، می تواند متفاوت باشد! و این واقعیتی است که نه تنها برای درخت های فیلوژنتیک مربوط به « پلاسمودیوم ها »، بلکه برای ژن های متفاوت موجودات زنده ی دیگر اعم از دلفین، خفاش، اسفنج، انسان و … نیز به چشم می خورد که ان شاء الله در مقالات آتی که مفصلاً به بحث « ژنتیک و تکامل » مربوط می شود، به مواردی از آن ها اشاره خواهیم کرد!
بدین ترتیب مشخص می گردد که در بررسی های ژنتیکی و مطالعات مبتنی بر DNA، درخت های فیلوژنتیک مربوط به بررسی های خویشاوندی و تکاملی، همواره نتایج یکسان و حتی مشابهی به دست نمی دهند! بلکه بستگی به این که کدام قطعه از DNA یا کدام ژن موجودات زنده، با یکدیگر مقایسه شود، نتایج متفاوت و درخت های فیلوژنتیک گوناگونی حاصل می گردد که این امر با ادعاهای تکامل شناسان، قرابت زیادی ندارد! چرا که اگر سخنان آن ها در مورد قرابت برخی از موجودات زنده با یکدیگر و دوری برخی از آن ها از یکدیگر درست باشد، اصولاً نباید تفاوت های زیادی در بررسی های ژنتیکی ژن های مختلف آن ها وجود داشته باشد! حال آن که در بسیاری از موارد، درخت های فیلوژنتیک رسم شده بر اساس ژن های مختلف چند موجود زنده ی « به اصطلاح هم خانواده »، تفاوت های فاحشی را به نمایش می گذارند!
این سخن تنها ادعایی از جانب ما نیست؛ بلکه مقالات و مطالعات متعدد علمی نیز به وجود چنین ناهماهنگی هایی به صراحت اشاره کرده و در صدد اختراع و ابداع روش های آنالیز، نرم افزار های کامپیوتری و ترفندهای آماری هستند تا بتوانند مشکلات ناشی از این ناهماهنگی ها را به حداقل برسانند!:(۲۰۵)
در بررسی های ژنتیکی و مطالعات مبتنی بر DNA، درخت های فیلوژنتیک مربوط به بررسی های خویشاوندی و تکاملی، همواره نتایج یکسان و حتی مشابهی به دست نمی دهند! بلکه بستگی به این که کدام قطعه از DNA یا کدام ژن موجودات زنده، با یکدیگر مقایسه شود، نتایج متفاوت و درخت های فیلوژنتیک گوناگونی حاصل می گردد که این امر با ادعاهای تکامل شناسان، قرابت زیادی ندارد! چرا که اگر سخنان آن ها در مورد قرابت برخی از موجودات زنده با یکدیگر و دوری برخی از آن ها از یکدیگر درست باشد، اصولاً نباید تفاوت های زیادی در بررسی های ژنتیکی ژن های مختلف آن ها وجود داشته باشد! حال آن که در بسیاری از موارد، درخت های فیلوژنتیک رسم شده بر اساس ژن های مختلف چند موجود زنده ی « به اصطلاح هم خانواده »، تفاوت های فاحشی را به نمایش می گذارند! این سخن تنها ادعایی از جانب ما نیست؛ بلکه مقالات و مطالعات متعدد علمی نیز به وجود چنین ناهماهنگی هایی به صراحت اشاره کرده و در صدد اختراع و ابداع روش های آنالیز، نرم افزار های کامپیوتری و ترفندهای آماری هستند تا بتوانند مشکلات ناشی از این ناهماهنگی ها را به حداقل برسانند!
در بررسی های ژنتیکی و مطالعات مبتنی بر DNA، درخت های فیلوژنتیک مربوط به بررسی های خویشاوندی و تکاملی، همواره نتایج یکسان و حتی مشابهی به دست نمی دهند! بلکه بستگی به این که کدام قطعه از DNA یا کدام ژن موجودات زنده، با یکدیگر مقایسه شود، نتایج متفاوت و درخت های فیلوژنتیک گوناگونی حاصل می گردد که این امر با ادعاهای تکامل شناسان، قرابت زیادی ندارد! چرا که اگر سخنان آن ها در مورد قرابت برخی از موجودات زنده با یکدیگر و دوری برخی از آن ها از یکدیگر درست باشد، اصولاً نباید تفاوت های زیادی در بررسی های ژنتیکی ژن های مختلف آن ها وجود داشته باشد! حال آن که در بسیاری از موارد، درخت های فیلوژنتیک رسم شده بر اساس ژن های مختلف چند موجود زنده ی « به اصطلاح هم خانواده »، تفاوت های فاحشی را به نمایش می گذارند! این سخن تنها ادعایی از جانب ما نیست؛ بلکه مقالات و مطالعات متعدد علمی نیز به وجود چنین ناهماهنگی هایی به صراحت اشاره کرده و در صدد اختراع و ابداع روش های آنالیز، نرم افزار های کامپیوتری و ترفندهای آماری هستند تا بتوانند مشکلات ناشی از این ناهماهنگی ها را به حداقل برسانند!
بدین ترتیب همان گونه که ملاحظه فرمودید، مطالعات ژنتیکی بر روی ژن های مختلف موجودات زنده، نه تنها نتوانسته معمای خویشاوندی و ارتباط تکاملی موجودات زنده را حل کند، بلکه سوالات، ابهامات و پیچیدگی های زیادی را به همراه داشته است!
این سوالات، ابهامات و پیچیدگی ها، موجب شده است تا بعد از فراگیر شدن استفاده از کامپیوتر و نرم افزارهای کامپیوتری، زیست شناسان و تکامل شناسان از نرم افزارها و برنامه های کامپیوتری متعددی برای تلفیق و مرتبط کردن الگوریتم های ناهمگون بهره ببیند تا شاید بتوانند مشکلات ناشی از ناهماهنگی ها و تناقضات مشاهده شده در آنالیز ژن های مختلف را به نحوی کاهش دهند!(۲۰۵) اما هنوز برنامه ای که بتواند بر این ناهمگونی ها غلبه کند، ابداع نشده است!(۲۰۵) همچنین برخی از محققین با مشاهده ی ناتوانی الگوریتم های مختلف در متحد ساختن اطلاعات حاصل از آنالیز های ژنتیکی، عنوان کرده اند که اصولاً ممکن است نتوان داده های حاصل از آنالیزهای ژنتیکی را در درخت های فیلوژنتیک که کامل ترین آن ها به عنوان « درخت فیلوژنتیک زندگی : Phylogenetic Tree of Life » نامیده می شود، قرار داد!(۲۰۵)
البته چنین نتایجی کاملاً قابل انتظار است! اصولاً تا زمانی که تمامی موجودات زنده ی کره ی زمین کشف نشوند، نمی توان الگوریتم مناسبی که الگوهای ژنتیکی موجودات زنده ی مختلف را پوشش دهد، طراحی نمود!
به هر حال ناسازگاری های موجود بین نتایج حاصل از مطالعات ژنتیکی و درخت های فیلوژنتیک مبتنی بر DNA موجودات زنده، چالشی بزرگ برای زیست شناسان و به خصوص تکامل شناسان به شمار می رود؛ به عبارت دیگر، گرچه قیافه ی حق به جانب تکامل شناسان و ادعای آن ها در مورد آنالیزهای ژنتیکی شان، ممکن است مخاطبان (به خصوص مخاطبان عام) را فریب دهد، اما این مسئله، تضادها، بحث ها و چالش های جدی در بین دانشمندان به جریان انداخته است که هنوز تا رفع آن ها، فاصله ی زیادی وجود دارد!
البته « تکامل شناسان » که دروغ، دغل و فریبکاری بخش بزرگی از حیاتشان را تشکیل می دهد، معمولاً با ارایه ی درخت های فیلوژنتیک هدف دار، سعی می کنند تا سایر درخت های فیلوژنتیک را که با ادعاهایشان تناقض دارد، سانسور نموده و به لطایف الحیل، از چشم مخاطبان خود دور بدارند! در واقع آن ها در مقالات خود که در دفاع از « فرضیه ی تکامل » می نگارند، فقط به ارایه ی درخت های فیلوژنتیکی مبادرت می ورزند که نشان می دهد ادعاهای آن ها درست می باشد! و درخت های فیلوژنتیک متناقض یا ناهماهنگ با ادعاهایشان را سانسور می نمایند! برای درک بهتر این مسئله، مجدداً مثال مربوط به مقایسه ی ژن های « پلاسمودیوم ها » را از این زاویه نیز بررسی می نماییم:
فرض می کنیم که تکامل شناسان بخواهند اثبات نمایند که انگل های دو گونه ی « پلاسمودیوم الونگاتوم : P. elongatum » و « پلاسمودیوم گالیناسئوم : P. gallinaceum »، از نظر تکاملی و ژنتیکی به یکدیگر نزدیک هستند! آن ها برای اثبات این ادعای خودشان، در مقالات مربوط به فرضیه ی تکامل، فقط آنالیز به دست آمده از « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » را که مطابق ادعاهایشان بوده و شباهت ژنتیکی دو گونه ی « پلاسمودیوم الونگاتوم : P. elongatum » و « پلاسمودیوم گالیناسئوم : P. gallinaceum » را به نمایش می گذارد، به مخاطبان خود عرضه می کنند!
به عبارت دیگر، آن ها با زیرکی تمام، تنها درخت فیلوژنتیک به دست آمده از « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » را که مطابق ادعاهایشان است، ارایه می کنند؛ حال آن که درخت های فیلوژنتیک ناشی از آنالیز بخش های دیگر DNA و ژن های دیگر « پلاسمودیوم ها » را که با ادعاهایشان تناقض و ناهماهنگی دارد، پنهان می کنند! بدین ترتیب که آنالیز « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » را که با ادعاهایشان ناهماهنگ بوده و دو گونه ی « پلاسمودیوم الونگاتوم : P. elongatum » و « پلاسمودیوم گالیناسئوم : P. gallinaceum » را به نسبت دور از هم نشان می دهد، در کتب و مقالاتشان به نمایش نمی گذارند!:
فرض می کنیم که تکامل شناسان بخواهند اثبات نمایند که انگل های دو گونه ی « پلاسمودیوم الونگاتوم : P. elongatum » و « پلاسمودیوم گالیناسئوم : P. gallinaceum »، از نظر تکاملی و ژنتیکی به یکدیگر نزدیک هستند! آن ها برای اثبات این ادعای خودشان، در مقالات مربوط به فرضیه ی تکامل، فقط آنالیز به دست آمده از « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » (درخت وسط) را که مطابق ادعاهایشان بوده و شباهت ژنتیکی دو گونه ی « پلاسمودیوم الونگاتوم : P. elongatum » و « پلاسمودیوم گالیناسئوم : P. gallinaceum » را به نمایش می گذارد، به مخاطبان خود عرضه می کنند! به عبارت دیگر، آن ها با زیرکی تمام، تنها درخت فیلوژنتیک به دست آمده از « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » را که مطابق ادعاهایشان است، ارایه می کنند (درخت وسط)؛ حال آن که درخت های فیلوژنتیک ناشی از آنالیز بخش های دیگر DNA و ژن های دیگر « پلاسمودیوم ها » را که با ادعاهایشان تناقض و ناهماهنگی دارد، پنهان می کنند! بدین ترتیب که آنالیز « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » را که با ادعاهایشان ناهماهنگ بوده و دو گونه ی « پلاسمودیوم الونگاتوم : P. elongatum » و « پلاسمودیوم گالیناسئوم : P. gallinaceum » را به نسبت دور از هم نشان می دهد (درخت سمت چپ)، در کتب و مقالاتشان مخفی می نمایند!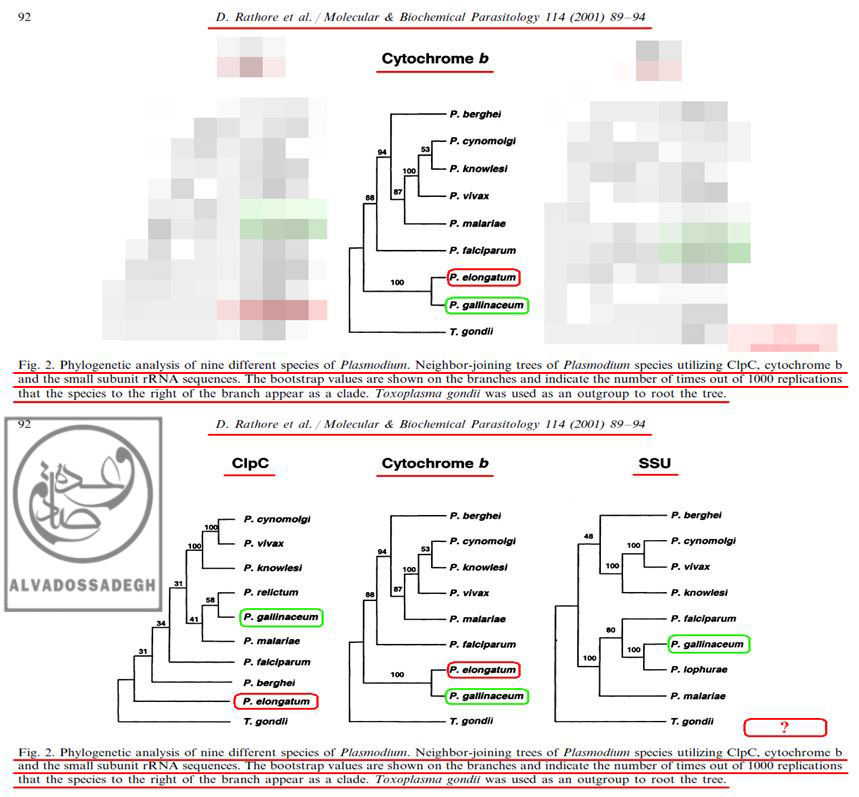
در مثالی دیگر، فرض می کنیم که تکامل شناسان تلاش می کنند تا به مخاطبان خود اثبات نمایند که انگل های دو گونه « پلاسمودیوم گالیناسئوم : P. gallinaceum » و « پلاسمودیوم مالاریه : P. malariae »، از نظر تکاملی و ژنتیکی به یکدیگر نزدیک هستند!
آن ها برای اثبات این ادعای خودشان، در مقالات مربوط به فرضیه ی تکامل، فقط آنالیز به دست آمده از « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » را که مطابق ادعاهایشان بوده و شباهت ژنتیکی دو گونه ی « پلاسمودیوم گالیناسئوم : P. gallinaceum » و « پلاسمودیوم مالاریه : P. malariae » را به نمایش می گذارد، به مخاطبان خود عرضه می کنند!
به عبارت دیگر، آن ها با زیرکی تمام، تنها درخت فیلوژنتیک به دست آمده از « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » را که مطابق ادعاهایشان است، ارایه می کنند؛ حال آن که درخت های فیلوژنتیک ناشی از آنالیز بخش های دیگر DNA و ژن های دیگر « پلاسمودیوم ها » را که با ادعاهایشان تناقض و ناهماهنگی دارد، پنهان می نمایند!
بدین ترتیب که آنالیز « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » و نیز آنالیز « DNA هسته ای کد کننده ی زیر واحد کوچک RNA ریبوزومی (rRNA) موسوم به (SSU) » را که با ادعاهایشان ناهماهنگ بوده و دو گونه ی « پلاسمودیوم گالیناسئوم : P. gallinaceum » و « پلاسمودیوم مالاریه : P. malariae » را به نسبت دور از هم نشان می دهد، در کتب و مقالاتشان به نمایش نمی گذارند و از دید اکثر مخاطبان (به خصوص مخاطبان عام)، مخفی می نمایند!:
فرض می کنیم که تکامل شناسان بخواهند اثبات نمایند که انگل های دو گونه « پلاسمودیوم گالیناسئوم : P. gallinaceum » و « پلاسمودیوم مالاریه : P. malariae »، از نظر تکاملی و ژنتیکی به یکدیگر نزدیک هستند! آن ها برای اثبات این ادعای خودشان، در مقالات مربوط به فرضیه ی تکامل، فقط آنالیز به دست آمده از « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » (درخت سمت چپ) را که مطابق ادعاهایشان بوده و شباهت ژنتیکی دو گونه ی « پلاسمودیوم گالیناسئوم : P. gallinaceum » و « پلاسمودیوم مالاریه : P. malariae » را به نمایش می گذارد، به مخاطبان خود عرضه می کنند! به عبارت دیگر، آن ها با زیرکی تمام، تنها درخت فیلوژنتیک به دست آمده از « DNA پلاستیدی کد کننده ی پروتئاز کازئینولیتیک موسوم به (ClpC) » را که مطابق ادعاهایشان است، ارایه می کنند (درخت سمت چپ)؛ حال آن که درخت های فیلوژنتیک منبعث از آنالیز بخش های دیگر DNA و ژن های دیگر « پلاسمودیوم ها » را که با ادعاهایشان تناقض و ناهماهنگی دارد، پنهان می نمایند! بدین ترتیب که آنالیز « DNA میتوکندریال (mtDNA) کد کننده ی پروتئین سیتوکروم b موسوم به (Cytochrome b) » (درخت وسط) و نیز آنالیز « DNA هسته ای کد کننده ی زیر واحد کوچک RNA ریبوزومی (rRNA) موسوم به (SSU) » (درخت سمت راست) را که با ادعاهایشان ناهماهنگ بوده و دو گونه ی « پلاسمودیوم گالیناسئوم : P. gallinaceum » و « پلاسمودیوم مالاریه : P. malariae » را به نسبت دور از هم نشان می دهد، در کتب و مقالاتشان به نمایش نمی گذارند!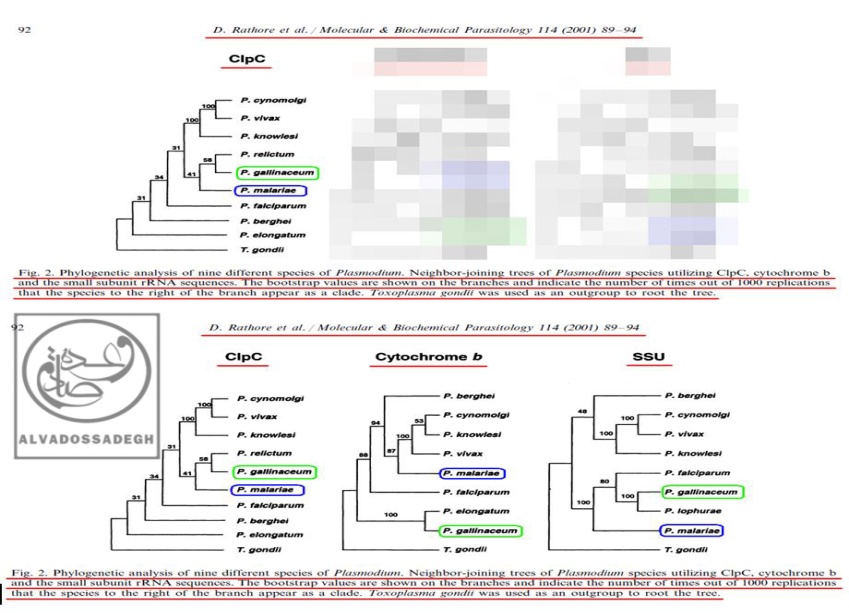
در واقع تکامل شناسان همچون ماهی لغزنده ای که از دست ماهیگیر فرار می کند و از دست صیاد می گریزد، تلاش می کنند تا با چنین حربه هایی از دست منتقدان احتمالی بگریزند و شواهد علمی که ممکن است از سوی منتقدان علیه آن ها استفاده شود را تا حد امکان در بایکوت رسانه ای قرار دهند! ان شاء الله در سلسله مقالات آتی، مثال های عملی و عینی چنین فریبکاری هایی را خدمت مخاطبان محترم، ارایه خواهیم نمود!
البته چنین تاکتیک هایی دیگر سودی برای آن ها نخواهد داشت! چرا که با رشد علمی جوامع و نیز دسترسی بیشتر دانشمندان به منابع مختلف، پنهان کاری های علمی، بسیار سخت تر از قبل خواهد بود!
با توجه به مطالبی که تاکنون بیان شد، می توان دریافت که گرچه مطالعه ی فسیل ها با استفاده از آنالیز ژنی مبتنی بر پروتکل « DNA باستانی : Ancient DNA »، دقیق تر از مطالعه بر اساس آناتومی و ظاهر فسیل ها می باشد، اما این روش نیز علی رغم ظاهر جذاب، فریبنده و دهان پرکنی که دارد، با ایرادات، اشکالات و ابهامات بی شماری مواجه است که اعتماد به آن را نیز غیر ممکن می کند!
ما در صفحات قبل، موارد متعددی از ضعف ها و ایرادات مربوط به مطالعه ی فسیل ها بر مبنای روش « DNA باستانی : Ancient DNA » را برشمردیم و مطالعات بیشتر در این زمینه را به مخاطبان محترم واگذار می کنیم. البته این ضعف ها و اشکالات تنها توهم! ما نیست و محققان دیگر نیز به اشکالات این روش، اشاره کرده اند و حتی دانشمندان و دانش پژوهان را به دقت بیشتر در این گونه اشکالات، حفظ روحیه ی انتقادی در مورد این گونه مطالعات و نیز کنکاش جدی در مورد باورپذیری، صحت و قابل اطمینان بودن نتایج حاصل از این تحقیقات، دعوت نموده اند!:
بسیاری از محققان، به اشکالات موجود در مطالعه ی فسیل ها به روش « DNA باستانی : Ancient DNA » ، اشاره کرده اند و حتی دانشمندان و دانش پژوهان را به دقت بیشتر در این گونه اشکالات، حفظ روحیه ی انتقادی در مورد این گونه مطالعات و نیز کنکاش جدی در مورد باورپذیری، صحت و قابل اطمینان بودن نتایج حاصل از این تحقیقات، دعوت نموده اند!
اما علی رغم مسایلی که گفته شد، مشکل بزرگی دامن گیر جامعه ی علمی کشورمان شده است که همانا عدم وجود اعتماد به نفس و عدم پرورش روحیه ی انتقادی در بسیاری از دانش پژوهان و دانشمندان این مرز و بوم می باشد! این امر به خصوص در مواردی که اصطلاحات جذاب و فریبنده ای همچون « ژنتیک »، « DNA »، « پروتئومیکس »، « بیوتکنولوژی »، « نانوتکنولوژی »، « سلول های بنیادی » و … از سوی دانشمندان غربی عنوان می شود، بیش از پیش خود را نشان می دهد! به نحوی که جامعه ی آکادمیک کشورمان که سرشار از دانشجویان و اساتید با هوش و با سواد نیز می باشد، در مقابل این اصطلاحات و ادعاهای دانشمندان غربی در این زمینه ها، تسلیم شده و بسیاری از ادعاهای آن ها را چشم بسته و بدون تفکر می پذیرد!
البته این ایراد بیمارگونه مختص جامعه ی علمی ما نیست! بلکه مشابه همین روند، با شدتی کمتر در جوامع غربی نیز مشاهده می گردد! به نحوی که برخی از دانشمندان متفکر و دغدغه مند نیز زبان به اعتراض در این زمینه گشوده و جامعه ی علمی را از اعتماد بی محابا و بدون تفکر نسبت به متدهای جدید « مهندسی ژنتیک » و مطالعات مبتنی بر چنین روش هایی، بر حذر داشته و خواهان حاکمیت مجدد تفکر بر جوامع علمی شده اند!
برای مثال، در مقاله ی زیر که در نقد پروژه ی بین المللی معروف ژنتیکی موسوم به « HapMap » نگاشته شده و در سال ۲۰۰۶ میلادی در نشریه ی معروف « مجله ی اروپایی ژنتیک انسانی : European Journal of Human Genetics » وابسته به انتشارات مشهور « Nature Publishing Group » منتشر شده است، مولفان مقاله، به صراحت، ادعاها و سر و صداهای زیادی که در مورد پروژه ی ژنتیکی بین المللی « HapMap » به راه افتاده است را زیر سوال برده و از حمله ی فله ای! و گروهی! دانشمندان به سمت « مطالعات ژنتیکی »، آن هم بدون این که پشتوانه ی فکری دقیق و عالمانه ای، زمینه ساز بسیاری از این مطالعات باشد، به شدت ابراز ناراحتی و گلایه نموده اند! به نحوی که در بخش انتهایی این مقاله، چنین مطلبی را درج نموده اند:(۲۰۷)
»It would be far better to spend more time thinking and planning before jumping in to genotyping every sample we can get our hands on. «
« بسیار بهتر خواهد بود که ما وقت بیشتری را به تفکر و برنامه ریزی اختصاص دهیم، قبل از این که هر نمونه ای که به دستمان برسد را فوراً آنالیز ژنتیکی نماییم! »(۲۰۷)
دانشمندان متفکر و دغدغه مند نیز، زبان به اعتراض گشوده و جامعه ی علمی را از اعتماد بی محابا و بدون تفکر نسبت به متدهای جدید « مهندسی ژنتیک » و مطالعات مبتنی بر چنین روش هایی، بر حذر داشته و خواهان حاکمیت مجدد تفکر بر جوامع علمی شده اند! برای مثال، در مقاله ی فوق که در نقد پروژه ی بین المللی معروف ژنتیکی موسوم به « HapMap » نگاشته شده و در سال ۲۰۰۶ میلادی در نشریه ی معروف « مجله ی اروپایی ژنتیک انسانی : European Journal of Human Genetics » وابسته به انتشارات مشهور « Nature Publishing Group » منتشر شده است، مولفان مقاله، به صراحت، ادعاها و سر و صداهای زیادی که در مورد پروژه ی ژنتیکی بین المللی « HapMap » به راه افتاده است را زیر سوال برده و از حمله ی فله ای! و گروهی! دانشمندان به سمت « مطالعات ژنتیکی »، آن هم بدون این که پشتوانه ی فکری دقیق و عالمانه ای، زمینه ساز بسیاری از این مطالعات باشد، به شدت ابراز ناراحتی و گلایه نموده اند! به نحوی که در بخش انتهایی این مقاله، چنین مطلبی را درج نموده اند: « بسیار بهتر خواهد بود که ما وقت بیشتری را به تفکر و برنامه ریزی اختصاص دهیم، قبل از این که هر نمونه ای که به دستمان برسد را فوراً آنالیز ژنتیکی نماییم! »
چنین انتقاداتی آن هم نسبت به پروزه های مشهوری هم چون « HapMap » که ارتباط مستقیمی هم با مقوله ی « تکامل » ندارد و بیشتر در حیطه ی ژنتیک پزشکی مطرح می گردد و تعریف ها و تمجیدهای بی شمار کورکورانه ای از سراسر جهان (حتی از کشور ما!) را نیز نسبت به خود بر می انگیزد، یک درس بسیار خوب برای ما دارد: « بسیاری از پروژه های مشهور علمی، متد پیشرفته، اما تفکر و برنامه ی ضعیفی دارند! و متاسفانه این مطالعات، با اصطلاحات دهان پرکن و روش های جدیدشان، تلاش می کنند تا عقبه ی ضعیف فکری و خلاء برنامه ریزی دقیق خود را بپوشانند! ».
ما دانشمندان و دانش پژوهان ایران زمین، نباید تنها نظاره گر باشیم! هنگامی که اصطلاحات فریبنده و جذابی چون « مطالعه ی ژنتیک »، مطالعه بر روی « DNA »، مطالعه به روش « DNA باستانی : Ancient DNA » و … را می شنویم، نباید تصور کنیم که استفاده از این روش ها در مطالعات، به معنای دقت و صحت نتایج این مطالعات است! همچنین نباید تصور کنیم که ادعای محققان مذکور درباره ی نتایج مطالعات، واقعاً و حقیقتاً با آن چه که مطالعه در عمل ارایه می کند، مترادف است! چه بسا مطالعات متعدد ژنتیکی با روش های پیشرفته انجام می شوند، اما به دلیل ضعف عقبه ی فکری، نتایجی که به دست می دهند، ارزشی ندارد!
به هر حال با توجه به مطالبی که گفته شد، نتیجه می گیریم که مطالعه ی فسیل ها به روش « DNA باستانی : Ancient DNA »، گرچه از روش مطالعه ی ظاهری فسیل ها، بهتر و منطقی تر می باشد، ولی این روش نیز با مشکلات، چالش ها، ابهامات و ضعف های متعددی رو به رو است و به نتایج حاصل از آن نیز نمی توان اعتماد کرد! بنابراین به این روش باید به مثابه کودک نوزادی نگاه کرد که راهی بسیار طولانی تا بالغ شدن و قابل اعتماد گردیدن در پیش دارد!
ادامه دارد …
خادم الامام (عج) – وعده صادق
بخش بعد: آیا ادعاهای موجود پیرامون اطلاعات کسب شده ازنئاندرتا ها صحیح است؟… DNA
منابع و مآخذ
Green, R. E., et al. (2008). A complete Neandertal mitochondrial genome sequence determined by high-throughput sequencing. Cell, 134, 416–۴۲۶٫
Krings, M; Stone, A; Schmitz, R W; Krainitzki, H; Stoneking, M; Paabo, S. Neandertal DNA sequences and the origin of modern humans. Cell 90 (1997): 19-30.
Noonan JP, Coop G, Kudaravalli S, Smith D, Krause J, et al. (2006) Sequencing and analysis of Neanderthal genomic DNA. Science 314: 1113-1118.
Green RE, Krause J, Ptak SE, Briggs AW, Ronan MT, et al. (2006) Analysis of one million base pairs of Neanderthal DNA. Nature 16: 330-336.
Ankel-Simons F, Cummins JM. Misconceptions about mitochondria and mammalian fertilization: implications for theories on human evolution. ProcNatlAcadSci U S A. 1996 Nov 26; 93(24):13859-63.
Schwartz M, Vissing J (2002). Paternal inheritance of mitochondrial DNA. N Engl J Med 347: 576–۵۸۰٫
Gibbons A (January 1998), “Calibrating the mitochondrial clock”, Science 279 (5347): 28–۹٫
http://www.dnai.org/teacherguide/pdf/reference_romanovs.pdf
Tully, L.A. et al., A sensitive denaturing gradient-gel electrophoresis assay reveals a high frequency of heteroplasmy in hypervariable region 1 of the human mitochondrial DNA control region. Am. J. Hum. Genet., 2000; 67: 432-443.
Calloway CD, Reynolds RL, Herrin GL, Anderson WW: The frequency of heteroplasmy in the HVII region of mtDNA differs across tissue types and increases with age. Am J Hum Genet 2000, 66(4):1384-1397.
http://ocw.tufts.edu/Content/20/CourseHome/301011/301031
http://www.newcastle-mitochondria.com/mitochondria/what-do-mitochondria-do/
Calloway CD, Reynolds RL, Herrin GL, Anderson WW: The frequency of heteroplasmy in the HVII region of mtDNA differs across tissue types and increases with age. Am J Hum Genet 2000, 66(4):1384-1397.
Sondheimer N, Glatz CE, Tirone JE, Deardorff MA, Krieger AM, Hakonarson H: Neutral mitochondrial heteroplasmy and the influence of aging. Hum Mol Genet 2011, 20(8):1653-1659.
https://www.promega.com/~/media/files/resources/conference%20proceedings/ishi%2010/oral%20presentations/35calloway.pdf?la=en
Gibbons A (January 1998), “Calibrating the mitochondrial clock”, Science 279 (5347): 28–۹٫
http://www.dnai.org/teacherguide/pdf/reference_romanovs.pdf
Howell N, Howell C, Elson JL (2008) Time dependency of molecular rate estimates for mtDNA: this is not the time for wishful thinking. Heredity, 101, 107–۱۰۸٫
Gibbons A (January 1998), “Calibrating the mitochondrial clock”, Science 279 (5347): 28–۹٫
http://www.dnai.org/teacherguide/pdf/reference_romanovs.pdf
Howell N, Howell C, Elson JL (2008) Time dependency of molecular rate estimates for mtDNA: this is not the time for wishful thinking. Heredity, 101, 107–۱۰۸٫
White DJ, Wolff JN, Pierson M and Gemmell NJ (2008) Revealing the hidden complexities of mtDNA inheritance. Molecular Ecology 17: 4925–۴۹۴۲٫
http://epidemic.bio.ed.ac.uk/how_to_read_a_phylogeny
http://guava.physics.uiuc.edu/~nigel/courses/598BIO/498BIOonline-essays/hw2/files/hw2_li.pdf
http://www.answers.com/topic/phylogenetic-tree
http://en.wikipedia.org/wiki/Phylogenetic_tree
Pearson WR, Robins G, Zhang T. Generalized neighbor-joining: more reliable phylogenetic tree reconstruction. MolBiolEvol. 1999 Jun;16(6):806-16.
http://www.icp.ucl.ac.be/~opperd/private/neighbor.html
http://www.answers.com/topic/neighbor-joining
http://en.wikipedia.org/wiki/Neighbor_joining
http://stat.duke.edu/people/theses/WuY.html
http://www.answers.com/topic/bayesian-inference-in-phylogeny
http://en.wikipedia.org/wiki/Bayesian_inference_in_phylogeny
Rathore, D., A. M. Wahl, M. Sullivan, and T. F. McCutchan. 2001. A phylogenetic comparison of gene trees constructed from plastid mitochondrial and genomic DNA of Plasmodium species. Mol. Biochem. Parasitol. 114:89-94.
Jeffrey R, Eric CR (2007) Review of Phylogenetic Tree Construction University
of Louisville Bioinformatics Laboratory Technical Report Series 1-7.
Bapteste, E., E. Susko, J. Leigh, D. MacLeod, et al. 2005. “Do orthologous gene phylogenies really support tree-thinking?” Bmc Evolutionary Biology 5: 33-33.
Gilbert, T., Bandelt H.-J., Hofreiter, M. and Barnes I. (2005) Assessing ancient DNA studies. Trends in Ecology and Evolution, 20, 541-544.
Terwilliger JD, Hiekkalinna T (2006) An utter refutation of the “fundamental theorem of the HapMap”. Eur J Hum Genet 14: 426-437.
منبع : http://www.alvadossadegh.com/fa/vijhe-13/17113-217113.html


